Two-cell blastomere can remodel male gametes
Source:
Time: 2011-03-28
It is well known that mature oocytes can reprogram somatic nuclei in the process of nuclear transfer (NT) and remodel male gametes in normal fertilization. An intriguing question is whether blastomere from cleavage-stage embryos retains reprogramming activities. Egli et al reported that one blastomere of the two-cell-stage embryo can successfully reprogram a donor nucleus into an ES cell line or a chimeric mouse. Therefore, it is interesting to investigate whether two-cell blastomeres can remodel male gametes as mature oocytes do. Recently, a team of researchers, led by LI Jinsong, at the Institute of Biochemistry and Cell Biology, Shanghai Institutes for Biological Sciences, CAS, demonstrated that remodeling ability persists in the blastomeres of two-cell embryos.
To determine whether the two-cell blastomere can remodel male gametes to generate normal diploid embryos, YANG Hui and SHI Linyu, Ph.D. students in Dr. LI’s laboratory, produced haploid parthenogenetic two-cell blastomeres as recipients. They injected mature sperm or round spermatid into one blastomere of two-cell embryo, and observed that sperm heads could not be decondensed until blastomeres entered mitosis, while some of the round spermatids could form male nucleus before blastomere entry into mitosis. These observations suggest that both the cytoplasm and nucleus of two-cell blastomeres contain remodeling factors in which cytoplasmic factors alone are sufficient to remodel round spermatid male nucleus, but not sufficient to remodel the mature sperm, which also needs factors from blastomere nucleus. To generate blastocysts, in which, all cells contain genetic materials from both female and male donors, they injected one green-fluorescent round spermatid into each of the two blastomeres. Eight whole-green blastocysts were derived from 256 manipulated embryos. They cultured these blastocysts in the ES culture medium and generated four ES cell lines, which exhibited the same pluripotency as those of fertilization-derived ES cell lines. These results demonstrate that remodeling ability persists in the blastomeres of two-cell embryos.
This work entitled “Mice generated after round spermatid injection into haploid two-cell blastomeres” was published online in Cell Research on Mar 22nd, 2011.
This work was supported by the grants from the Chinese Academy of Sciences, the Ministry of Science and Technology, National Natural Science Foundation of China and Shanghai Municipal Commission for Science and Technology.
AUTHOR CONTACT:
LI Jinsong
Institute of Biochemistry and Cell Biology, Shanghai Institutes for Biological Sciences, Chinese Academy of Sciences, Shanghai, China
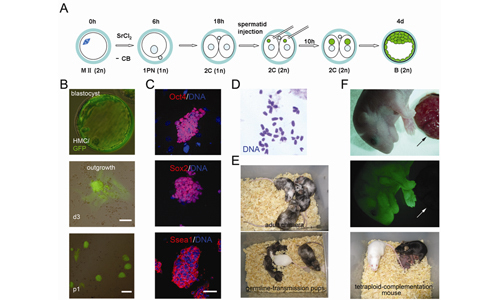
ES cells generated after ROSI into both blastomeres of two-cell haploid embryos.
(A) Experimental design. MII oocytes, activated in CB-free medium for 6 hr, form one pronucleus and develop to two-cell stage in vitro. Green-fluorescent round spermatid is injected into both blastomeres. Injected embryos develop to blastocysts, in which, all cells are green fluorescent.
(B) ES cells derived from green-fluorescent blastocyst. All cells are green fluorescent in blastocyst (top), outgrowth (middle, Scale bar, 50 μm) and ES cells (bottom, Scale bar, 100 μm).
(C) Expression of pluripotent markers in ES cells. Immunostaining with antibodies against Oct4 (top), Sox2 (middle) and Ssea1 (bottom). Counterstained with blue nuclear dye Hoechst 33342. Scale bar, 50 μm.
(D) Normal karyotype of ES cells.
(E) Healthy chimeric mouse generated from ES cells (top) and exhibited germline-transmission (bottom).
(F) Mouse produced by injection of ES cells into tetraploid blastocysts. Top, mouse and placenta, bright-field image. Middle, same mouse and placenta, fluorescent image. Mouse is green fluorescent. Arrow indicates the placenta, which is host-derived and thus not green fluorescent. Bottom, ES mouse (black) at 2 month with new-born pups. (Image provided by Dr. LI Jinsong)
 Appendix:
Appendix: